









Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkMotricidade
versão impressa ISSN 1646-107X
Motri. vol.11 no.2 Ribeira de Pena jun. 2015
https://doi.org/10.6063/motricidade.3385
ARTIGO DE REVISÃO
Exercício e mobilização de linfócitos T senescentes: Uma meta-análise
Exercise and mobilization of senescent T lymphocytes: A meta-analysis
Luciéle Guerra Minuzzi1,*; Ana Maria Miranda Botelho Teixeira1; José Pedro Ferreira1
1 Faculdade de Ciências do Desporto e Educação Física - Universidade de Coimbra, Coimbra, Portugal
RESUMO
O objectivo deste estudo foi avaliar a hipótese que o exercício físico induz aumentos na mobilização de células T senescentes. Foi realizado um levantamento sistematizado de estudos empíricos indexados nas bases de dados PUBMED e ScienceDirect, utilizando as palavras-chave: exercise, exercise-induced mobilization of T lymphocytes, exercise-induced mobilization of CD8, immune space, senescent T-cell mobilization, naive T-cell repertoire, senescent T-cell repertoire. Foram encontrados 442 estudos (PUBMED = 99 estudos; ScienceDirect = 343 estudos), sendo que seis preencheram todos os critérios definidos para inclusão na meta-análise. Os estudos que cumpriram os critérios de inclusão foram avaliados quanto a qualidade metodológica com a escala de PEDro e analisados utilizando o software Comprehensive Meta-analysis. Foram calculados a diferença entre os erros padrão da média (Std diff in means), o intervalo de confiança (95% CI), e os scores Z e P (Z-value e P-value). O exercício físico aumentou a mobilização de células T senescentes para o sangue periférico, quando comparados os valores agudos obtidos logo após o término do exercício com os valores pré-exercício. É possível que esta mobilização preferencial leve a indução da apoptose destas células criando espaço para a produção de células T virgens.
Palavras-chaves: Exercício, Células T senescentes, Imunosenescência, Espaço imune
ABSTRACT
The purpose of this study was to evaluate the hypothesis of increased mobilization of senescent T cells to the peripheral blood induced by exercise. The systematic survey of indexed empirical studies was conducted in PubMed and ScienceDirect databases using the keywords: exercise, exercise-induced mobilization of T lymphocytes, exercise-induced mobilization of CD8, immune space, senescent T-cell mobilization, naive T-cell repertoire, senescent T-cell repertoire. The main criterion was the mobilization of senescent T lymphocytes after exercise. After search, 442 studies (PUBMED = 99 studies; ScienceDirect = 343 studies) were found and six met all the criteria for inclusion in the meta- analysis. The studies that met the inclusion criteria were assessed for methodological quality with the PEDro scale. Data were analyzed using Comprehensive Meta-analysis software. The difference between the standard errors of the mean (Std diff in means), the confidence interval (95 % CI), and P and Z scores (Z-value and P-value) were calculated. In conclusion, acute exercise mobilizes a population of CD8+ T-cells exhibiting a senescent phenotype into the peripheral blood compartment. It is possible that this mobilization induces apoptosis of these cells and create a vacant space for the production of naive T cells.
Keywords: Exercise, senescent T cells, immunosenescence, immune space
INTRODUÇÃO
Teorias sobre o mecanismo do envelhecimento humano foram propostas, inúmeras vezes, ao longo dos últimos 2000 anos e ainda não temos uma explicação geralmente aceite e comprovada sobre este fenómeno universal da vida. Isto porque o envelhecimento consiste em vários eventos com uma variedade de causas, sendo que ninguém foi capaz de apontar uma causa predominante (Burzynski, 2003). Determinar até que ponto os fatores genéticos e o estilo de vida influenciam os declínios funcionais associados à idade e à magnitude das respostas adaptativas ao exercício (ou seja, treinabilidade) em populações mais jovens e mais idosas, é uma área de investigação ativa (Chodzko-Zajko et al., 2009).
Se o tempo de vida do ser humano aumenta, existe uma tendência a ocorrer, de modo concomitante, um aumento da carga de morbidade da população, levando a uma maior prevalência de doenças infeciosas e malignas relacionadas com a idade. A resistência de um organismo à doença e à infeção é comprometida com a idade devido ao aparecimento de declínios inevitáveis no funcionamento normal do sistema imune (Simpson et al., 2012). Durante o envelhecimento, em particular, a capacidade para combater infeções está diminuída pelo sistema imune, tal como evidenciado pelo aumento do número de infeções com sintomas mais graves, de duração prolongada e pior diagnóstico. Além disso, a reativação de infeções crónicas ocorre numa frequência maior em humanos e ratos idosos. Estas disfunções sugerem alterações em todos os componentes do sistema imune, mas as alterações mais consistentes e significativas são vistas no compartimento dos linfócitos T (Lages et al., 2008).
O braço adaptativo do sistema imune (isto é, células T, células B e seus produtos) parece diminuir mais com o aumento da idade e as alterações relacionadas com a idade no interior do compartimento das células T (isto é, a razão CD4/CD8 invertida, baixas respostas proliferativas, o aumento das células de memória e baixa síntese de IL-2) são características marcantes do Perfil de Risco Imune - uma mistura de biomarcadores imunes que foram utilizados para prever morbidade e mortalidade em idosos (Pawelec, 2006).
A expansão clonal de células T, em resposta a um estímulo antigénico é um processo fundamental da imunidade adaptativa, permitindo a identificação de antigénios específicos de células T efetoras para combater os patogénios invasores. No entanto, esta expansão clonal não é infinita, e após ciclos repetidos e excessivos de divisão celular, as células T sofrem paragem do ciclo celular e tornam-se senescentes. Neste estado, as células T não serão mais expandidas clonalmente sob futura estimulação antigénica, mas ainda mantém as propriedades de células efetoras (por exemplo, reconhecer e matar as células infetadas por vírus) e ainda são capazes de produzir grandes quantidades de citocinas pró-inflamatórias, tais como TNF-a, IFN-g, TNF-g (Spielmann et al., 2011). Nomeadamente, a estimulação antigénica faz com que células T proliferem (sofrem expansão clonal via divisão celular) e se diferenciem em células T efetoras que desempenham funções especializadas, tais como a secreção de citocinas, reconhecimento e morte de células alvo, e a ativação de macrófagos e células B produtoras de anticorpos. A exposição antigénica repetida (isto é, ao longo da vida) aumenta a frequência de células T senescentes no sangue e tecidos. O aparecimento de células T senescentes ocorre devido ao excesso de expansões clonais que ocorrem como parte de uma resposta imune normal à reativação viral ou à invasão do organismo ao longo da vida. Isso, juntamente com uma redução de células T naive recém-funcionais saindo do timo atrofiado, contribui para a diminuição do repertório de células T naive (isto é, reduzido espaço imune).
Assim, a exposição repetida a estímulos antigénicos de crescimento durante todo o tempo de vida (ou seja, reativação de infeções virais latentes) leva a mais rounds de divisão celular e senescência prematura. Na medida em que essas células T senescentes ainda retêm funções imediatas das células efetoras (ou seja, morte de células infetadas por vírus), e são altamente pró-inflamatórias, o seu acúmulo no sangue e nos tecidos pode também contribuir para um certo número de patologias associadas com a inflamação (Simpson & Guy, 2010).
Linfócitos T senescentes são células experientes em antigénio que expressam KLRG1 e/ou CD57 na superfície celular (Simpson et al., 2008). A expressão de KLRG1, como marcador de senescência replicativa, é dependente do número de divisões celulares seguindo a estimulação antigénica. Células T CD8+ expressando KLRG1 são células experientes em antigénios com uma capacidade diminuída para proliferar, mas ainda mantendo capacidades efetoras imediatas, tais como o reconhecimento e morte de células-alvo (Simpson et al., 2008). Linfócitos T CD8+ com alta expressão de CD57 tem uma reduzida habilidade de proliferar, citotoxicidade aumentada, produzem menos IL-2 mas mais IFN-g, e tem características indicativas de senescência replicativa, incluindo telômeros encurtados e aumento da expressão de KLRG1. Essas mesmas populações celulares falham em expressar CD28, uma importante molécula co-estimuladora para a ativação e proliferação de linfócitos T naive após um encontro antigénico.
Os efeitos benéficos do exercício podem vir tanto de perspetivas de prevenção e / ou rejuvenescimento (Simpson & Guy, 2010). Em primeiro lugar, a partir do ponto de vista da prevenção, o exercício pode provocar efeitos secundários sobre o sistema imune, devido às suas bem conhecidas propriedades redutoras de estresse, limitando assim o potencial de reativação viral latente induzida por estresse e erosão dos telómeros. Em segundo lugar, a partir do ponto de vista do tratamento/rejuvenescimento, o exercício regular pode levar à destruição do excesso de clones virais específicos de células T por meio de apoptose, libertando o ''espaço imune'' para ser ocupado pelas células T naive e expandindo o repertório antigénico de células T (Simpson, 2011). Para que este processo aconteça, Simpson and Guy (2010) propõem a existência de três fases distintas que devem ocorrer: 1) uma mobilização seletiva de células T senescentes dos tecidos periféricos para o compartimento do sangue durante o exercício; 2) extravasamento de células T senescentes da circulação e sua posterior apoptose nos tecidos periféricos durante a recuperação do exercício; 3) geração subsequente de células T naive para substituir as células T senescentes deletadas. Detalhadamente, o exercício agudo provoca a mobilização preferencial de células T altamente diferenciadas e senescentes dos tecidos periféricos para o compartimento do sangue (linfocitose), sob a influência de catecolaminas.
Por outro lado, o exercício aumenta a produção de espécies reativas de oxigénio (ROS), glicocorticóides, e citocinas pró-inflamatórias, expondo assim as células T senescentes a um meio de estímulos pró-apoptóticos. Recetores de superfície celular de morte (FAS / FAS ligante (FasL)) são up-regulados em células T senescentes, que também estão sujeitos a danos oxidativos de ADN no sangue. Estas células suscetíveis à apoptose, juntamente com células T naive e de memória danificadas, saem do compartimento sanguíneo durante a fase de recuperação do exercício (linfocitopenia) e migram para tecidos específicos. Uma parte destas células T senescentes subsequentemente sofrem apoptose nos tecidos periféricos, criando assim espaço vago. Por conseguinte, a redução no número de células T conduz a um feedback positivo, aumentando a saída de células T naive do timo ou de locais de desenvolvimento de células T extra timo (isto é, fígado, intestinos). Estas células T recém-geradas preenchem o espaço vago e contribuem para um repertório expandido de células T naive. Repetições deste processo em resposta ao exercício habitual reduzem o número de células T senescentes ao longo do tempo, diminuindo o risco de infeção e aumentando a longevidade saudável (Simpson & Guy, 2010).
No presente estudo realizamos uma pesquisa bibliográfica com o objetivo de avaliar a hipótese que o exercício físico induz aumentos na mobilização de células T senescentes.
MÉTODO
O levantamento bibliográfico sistematizado de estudos empíricos foi realizado nas bases de dados PUBMED e Science Direct, sendo o período selecionado para a busca o compreendido entre Janeiro de 2000 e Dezembro de 2012. Para a realização da busca on-line utilizaram-se os seguintes termos de busca ou descritores: exercise, exercise-induced mobilization of T lymphocytes, exercise-induced mobilization of CD8, immune space, senescent T-cell mobilization, naive T-cell repertoire, senescent T-cell repertoire.
Critérios de inclusão e exclusão na revisão
Na presente meta-análise foram incluídos apenas estudos publicados em língua inglesa. A pesquisa foi limitada a artigos de revistas científicas, não tendo sido consideradas teses e outras dissertações, capítulos de livros e estudos não publicados. A base de dados analisada também não incluiu artigos de revisão e estudos publicados antes do ano 2000.
O principal critério de inclusão foi a mobilização de linfócitos T senescentes após o exercício.
O interesse desta meta-análise era obter informações quanto aos efeitos agudos do exercício, pelo que foram excluídos os resultados de estudos com intervenções de suplementação e/ou treino, sendo apenas considerados os resultados dos efeitos agudos apresentados pelos diferentes estudos. Quando diferentes intensidades de exercício foram avaliadas, apenas foi incluída para efeito de análise, o valor de intensidade mais elevado. Quando diferentes faixas etárias foram consideradas, foram apenas incluídos os resultados referentes a indivíduos até 40 anos de idade. Não foi objetivo do estudo considerar a intensidade e a idade como variáveis moderadoras, podendo tal ser equacionado em futuros estudos.
Foram considerados apenas os linfócitos T CD8+ senescentes. Para determinar a senescência foram aceites os marcadores: KLRG1+, CD57+, KLRG1+/CD57+, KLRG1+/CD28-.
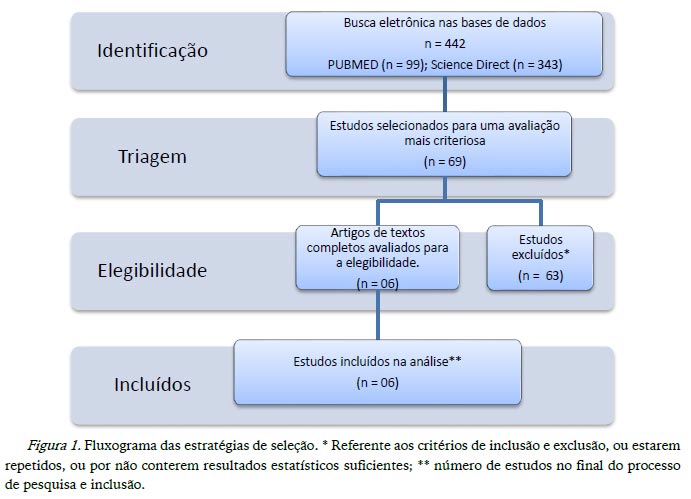
No presente estudo foram encontrados 442 artigos em duas bases de dados: PUBMED (99 estudos) e Science Direct (343 estudos). Após a identificação dos critérios de interesse através da leitura criteriosa dos respetivos resumos, foram selecionados um total de 69 estudos. Destes, seis preencheram todos os critérios definidos para inclusão na meta-análise. Os demais estudos (63) foram excluídos por: i) estarem repetidos nas diferentes base de dados e nos critérios de busca (n = 50), ii) serem artigos de revisão (n = 02), iii) não apresentarem resultados estatísticos que possibilitassem a análise do estudo ou iv) por não contemplarem os critérios de inclusão nomeados anteriormente (marcadores de senescência, por exemplo) (n = 11).
Análise Estatística
Depois de concluída a seleção dos estudos, os dados foram inseridos e projetados em gráficos do tipo forest plots e funnel plot com o auxílio de um software informático específico, o Comprehensive Meta-Analysis, version 2.0 copyright 2006.
Os dados foram classificados como sendo contínuos, tendo sido extraídos os valores de média e de erro padrão da média bem como o número total de indivíduos pertencentes a cada um dos estudos selecionados.
Para medir o tamanho do efeito (effect size) foi utilizado um intervalo de confiança (CI) de 95%. Os resultados foram inseridos e apresentados de forma a avaliar também a heterogeneidade (Chi2 e p<0,05) e a inconsistência (I2) para cada conjunto de dados (Higgins, Thompson, Deeks, & Altman, 2003).
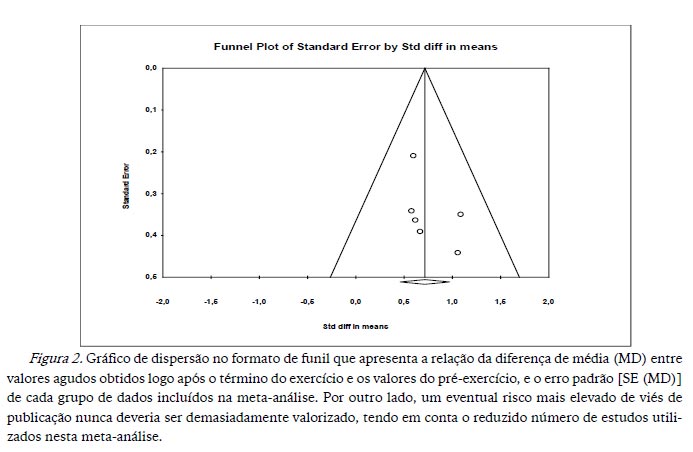
Para a seleção dos estudos, observou-se cuidadosamente possíveis vieses que pudessem prejudicar os resultados apresentados para cada um dos estudos. Foi utilizado o gráfico de dispersão no formato de funil para avaliar o risco de viés de publicação, por meio da avaliação da assimetria aplicada ao gráfico (Egger, Davey Smith, Schneider, & Minder, 1997). Quando não há qualquer risco de viés de publicação, o gráfico apresenta a figura de um funil invertido (Higgins & Green, 2008), sendo a simetria ou a assimetria definidas informalmente, por meio de avaliação visual (Egger et al., 1997).
Esta meta-análise foi realizada com base no modelo fixo.
A figura 1 apresenta um fluxograma que sintetiza as diferentes etapas e estratégias de seleção utilizadas, tendo por base a aplicação dos critérios de inclusão e exclusão.
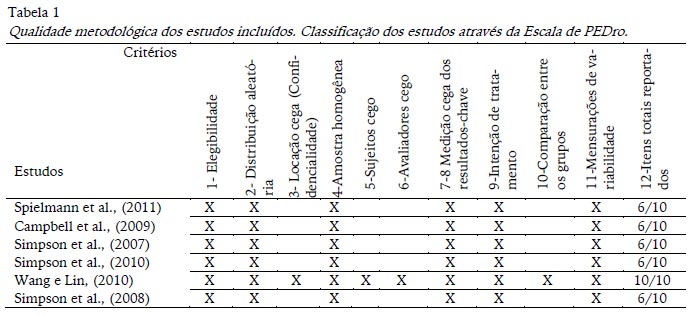
Os estudos que cumpriram os critérios de inclusão foram avaliados quanto à qualidade metodológica com a escala de PEDro, baseada na lista Delphi desenvolvida por Verhagen e colaboradores no Departamento de Epidemologia, da Universidade de Maastricht (Verhagen et al., 1998). Originalmente a escala de PEDro apresenta 11 critérios, no entanto os critérios número 7 e 8 (7. Todos os avaliadores que mediram pelo menos um resultado-chave, fizeram-no de forma cega; 8. Medições de pelo menos um resultado-chave foram obtidas em mais de 85% dos sujeitos inicialmente distribuídos pelos grupos) foram agrupados num único critério: Medição-cega dos resultados chave, porque nos pareceu mais apropriado. Se o score PEDro do estudo fosse menor que quatro, o estudo era classificado como de baixa qualidade metodológica e excluído. Nenhum dos seis estudos selecionados obteve pontuação inferior a quatro. A análise individual de cada estudo considerando a escala de PEDro está resumida na Tabela 1.
Depois de selecionados os estudos, os dados foram inseridos e analisados utilizando o software Comprehensive Meta-analysis. Foram calculados a diferença entre os erros padrões da média (Std diff in means), o intervalo de confiança (95% CI), e os scores Z e P (Z-value e P-value). Alguns procedimentos foram aplicados para corrigir viés causados por várias origens de erros, tais como erros da amostra e erros de medida.
RESULTADOS
Na análise foram incluídos seis estudos, que contabilizavam um total de 150 sujeitos. Quando avaliámos o risco de viés de publicação (Figura 2), por meio de assimetria, para cada conjunto de estudos sobre o exercício e a mobilização de linfócitos T senescentes, verificámos que os dados apresentavam um risco reduzido de viés de publicação, de acordo com os baixos valores de assimetria apresentados.
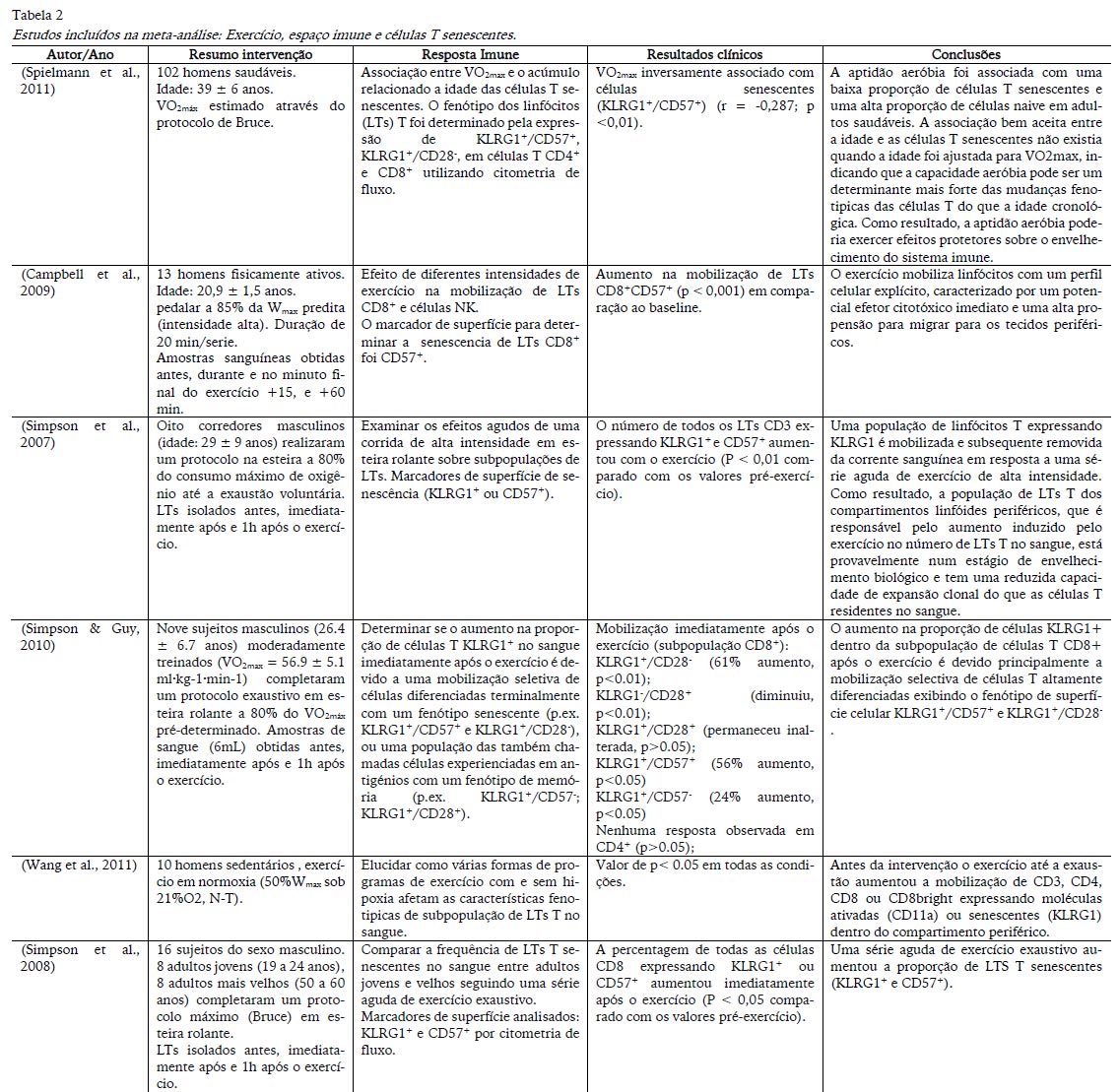
A tabela 2 apresenta os estudos selecionados para a meta-análise com autor/ano, resumo da intervenção (número de sujeitos, a média e o desvio padrão da idade, os testes utilizados e as variáveis analisadas), resposta imune (inclui os objetivos do estudo e as variáveis imune incluídas no estudo), conclusões (apresentação dos principais resultados e conclusões) e eficiência (apresenta o resumo estatístico dos principais resultados destacados nos estudos).
Todos os estudos investigaram sujeitos do sexo masculino. A idade compreendida variou de 20 a 40 anos e o nível de aptidão física alternou entre sedentários (Wang, Chen, & Weng, 2011), saudáveis (Campbell et al., 2009; Simpson et al., 2008; Spielmann et al., 2011) e treinados (Simpson et al., 2007, 2010).
Nos estudos de Campbell et al. (2009) e Wang, Chen, e Wen (2011) os sujeitos pedalaram em intensidades correspondentes a 85% e 50% da Wmáx predita, respetivamente. Nestes dois estudos os indivíduos não realizaram o esforço até a exaustão voluntária e apenas foram incluídos os resultados dos grupos que realizaram o exercício em normoxia (Wang et al., 2011) e em alta intensidade (Campbell et al., 2009). Os efeitos agudos foram observados após 20 minutos (Campbell et al., 2009) e 30 minutos (Wang et al., 2011) de ciclismo. Simpson et al. (2007) e Simpson et al. (2010) utilizaram o mesmo protocolo de exercício, corrida em passadeira rolante a 80% do VO2máx, enquanto Simpson et al. (2008) submeteu seus avaliados a um protocolo máximo em passadeira. No caso destes três estudos, os indivíduos realizaram o exercício até a exaustão. Por outro lado, Spielmann et al. (2011) correlacionou o VO2max com o acúmulo de células T senescentes. Neste caso, não foram consideradas as correlações relacionadas a idade que o estudo investigou.
Os estudos incluídos nesta meta-análise evidenciaram que linfócitos T com características senescentes são mobilizados dos compartimentos periféricos para o sangue imediamente após a realização de uma sessão de exercício. O padrão de aumento foi semelhante entre os estudos considerando o valor de p observado (Tabela 2).
Linfocitos T senescentes foram analisados quanto a presença dos marcadores de superfície celular KLRG1+, CD57+, CD28-. Somente o estudo de Campbell et al. (2009) não determinaram a senescência através desse marcador, mas através da expressão de CD57. O estudo de (Simpson et al., 2010) especialmente investigou se as células expressando KLRG1 eram linfócitos T terminalmente diferenciados (KLRG1+/CD28-/CD57+) ou uma população de células T memoria-efetoras (KLRG1+/CD28+/CD57-) que não atingiram a diferenciação terminal. Os resultados mostraram que o aumento na proporção de células KLRG1+ dentro da subserie de células T CD8+ após o exercício é devido principalmente a uma mobilização seletiva de células T altamente diferenciadas exibindo um fenótipo KLRG1+/CD57+ (56% aumento) e KLRG1+/CD28- (61% aumento) na superfície celular.
Diferente dos outros estudos que determinaram os efeitos agudos do exercício, Spielmann et al. (2011) examinaram a associação entre os valores de VO2max de homens saudáveis (18-24 anos) e o acúmulo de células T senescentes. Neste estudo somente foi considerado o grupo jovem. Análises de regressão univariada mostraram que o VO2máx foi inversamente associado com células T CD8+ senescentes (KLRG1+/CD57+, r = -0,287, p<0,01; KLRG1+/CD28-, r = -0,350, p<0,001), podendo indicar que a manutenção de altos níveis de aptidão aeróbia durante o curso natural do envelhecimento pode prevenir o acúmulo de células T senescentes com especificidade antigénica limitada.
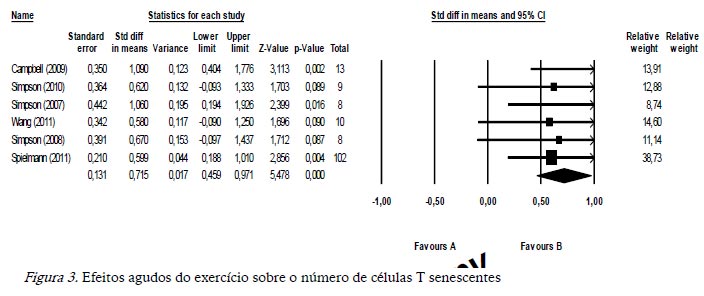
A figura 3 apresenta os resultados finais relativos aos 6 estudos incluídos no estudo, tendo por base a partir da utilização do programa Comprehensive Meta-analysis. O exercício físico aumentou o número de células T senescentes em todos os estudos analisados, ao comparar os valores agudos obtidos logo após o término do exercício com os valores pré-exercício. Os resultados são apresentados como a diferença entre os erros padrões da média (0,715), onde Z = 5,478 sendo esse efeito significativo (p < ,001).
DISCUSSÃO
O objetivo deste trabalho foi avaliar a hipótese do aumento da mobilização de células T senescentes induzido pelo exercício. A expansão de clones de células T CD8+ senescentes associadas a infeções virais persistentes ocupa uma grande proporção do espaço imune que, em condições de baixa exposição ao patogénio e reativações virais menos latentes, seria ocupado principalmente por células T naive. Na medida em que o número total de células T periféricas é estreitamente regulado, um acúmulo supérfluo de células T experientes em antigénio pode reduzir a saída de células T naive, diminuindo o número e a percentagem de células capazes de montar respostas imunes a novos agentes patogénicos. Nesta situação, estas células T resistentes à apoptose e altamente diferenciadas, sobrecarregam o espaço imune, traduzindo-se num menor repertório de células T para os antigénios novos, resultando numa vigilância imunológica reduzida e num risco aumentado de infeção (Simpson & Guy, 2010).
A análise dos seis estudos incluídos nesta meta-análise mostrou que o exercício físico ou uma boa aptidão aeróbia é capaz de aumentar mobilização de células T senescentes no sangue periférico. A mobilização de células T experientes em antigénio a partir dos tecidos periféricos para o sangue, poderia gerar um espaço vago onde as células T naive eventualmente ocupariam espaço (Simpson et al., 2007, 2008; Simpson & Guy, 2010). Essas células caracterizam-se por linfócitos citotóxicos com um perfil celular claramente definido, caracterizado por um imediato potencial citotóxico efetor e uma alta propensão para migrar dentro dos tecidos periféricos (Campbell et al., 2009).
Sugere-se que os linfócitos mobilizados com exercício sejam mais sensíveis a apoptose. Este processo poderia ser causado por estímulos pró-apoptóticos induzidos pelo exercício sobre as células T circulantes e provocando vias apoptóticas iniciais antes que a célula deixasse o compartimento do sangue. Como os linfócitos são conhecidos por saírem do sangue durante a fase de recuperação de exercício, uma parte destas células extravasadas (tendo já encontrado um estímulo pró-apoptótica) pode subsequentemente morrer por apoptose em tecidos periféricos (Simpson, 2011). Leucócitos CD57+ e KLRG1+ (senescentes) foram mais suscetíveis apoptose induzida por H2O2 in vitro comparado com linfócitos expressando CD28, CD62L, ou CD11a (Wang & Lin, 2010).
Os marcadores de senescência sobre as subpopulações de linfócitos T são considerados também eficientes marcadores do envelhecimento biológico sistémico (Simpson et al., 2008). Embora o CD45RA e o CCR7 sejam marcadores eficientes de diferenciação das células T, a identificação de células T CD45RA+/CCR7+ como células terminalmente diferenciadas (ex: senescentes) é controversa devido ao facto destas células ainda demonstrarem níveis eficazes de proliferação após ativação do recetor de célula T (TCR). Atualmente, a expressão combinada de KLRG1 e CD57, ou a expressão de KLRG1 na ausência de CD28 parecem ser marcadores mais aceites de senescência das células T, pois as células com este fenótipo são incapazes de sofrer futuras divisões, não expressam o CD27 e o CCR7, e têm uma baixa expressão de CD127 (Simpson, 2011; Simpson & Guy, 2010). Foi por esse motivo que esta meta-análise incluiu estudos que tivessem avaliado esta combinação de marcadores, embora essa opção tenha eliminado um certo número de estudos (11 no total), o que certamente é a maior limitação do estudo, procurou-se uma melhor confiabilidade e maior atualidade dos resultados.
Spielmann et al. (2011) descobriram que os indivíduos com valores de VO2máx acima da média tinham menos células T CD4+ e CD8+ senescentes e mais células T CD8+ naive do que aqueles com valores abaixo da média de VO2máx, mesmo após ajuste para idade, índice de massa corporal e percentagem de gordura corporal. Surpreendentemente, os autores verificaram que a associação, bem aceite, entre idade e o número de células T senescentes já não existia, quando a idade era ajustada para o VO2max, indicando que a aptidão aeróbia pode ser um forte determinante de mudanças fenotípicas de células T, mais do que a própria idade cronológica. Este efeito foi limitado às células senescentes, sendo que o VO2max não foi associado com as células de memória efetora (KLRG1+/CD28+) após o ajuste para a idade. Os autores consideraram que o seu estudo foi o primeiro a mostrar que a aptidão aeróbia está associada a uma moderação do acúmulo natural relacionado com a idade de células T senescentes no sangue periférico, com destaque para os efeitos benéficos da manutenção de um estilo de vida fisicamente ativo sobre o envelhecimento do sistema imunitário. Isso reforça a intuição de que a atividade física regular pode exercer propriedades preventivas e / ou de rejuvenescimento do envelhecimento do sistema imune por amenizar a acumulação relacionada com a idade, de células T senescentes e prevenir a rápida transição para a categoria de Perfil de Risco Imune mais tarde na vida (Simpson & Guy, 2010). Os autores postularam que a manutenção de altos níveis de aptidão aeróbia durante o curso natural do envelhecimento pode ajudar a prevenir o acúmulo de células T senescentes, que têm especificidade antigénica limitada, ao mesmo tempo, mantendo um número suficiente de células T naive capazes de reconhecer e responder a novos antigénios (Spielmann et al., 2011).
Os resultados do estudo de Spielmann et al. (2011) apontam uma série de implicações importantes para os benefícios preventivos do exercício contra a imunossenescência associada ao envelhecimento. Em primeiro lugar, dado que a proporção de células T senescentes aumenta em 10% por década de vida, na idade avançada, em homens saudáveis / assintomáticos, seria razoável postular que níveis elevados de aptidão física e exercício regular poderiam prevenir ou impedir o acúmulo de células T senescentes por décadas. Em segundo lugar, os níveis de atividade e aptidão física são os principais fatores moderadores na relação idade-imunossenescência e são preditores independentes da ''idade imunológica''. Em terceiro lugar, o valor terapêutico do exercício em melhorar a imunocompetência da população em envelhecimento assintomático e imunocomprometidos clinicamente (por exemplo, HIV +) é potencialmente imenso (Hong, 2011).
CONCLUSÕES
O exercício físico aumentou a mobilização de células T senescentes para o sangue periférico, quando comparados os valores agudos obtidos logo após o término do exercício com os valores pré-exercício. É possível que esta mobilização preferencial leve a indução da apoptose destas células criando espaço para a produção de células T virgens.
REFERÊNCIAS
Burzynski, S. R. (2003). Gene silencing--a new theory of aging. Medical Hypotheses, 60(4), 578–583. [ Links ]
Campbell, P. T., Campbell, K. L., Wener, M. H., Wood, B. L., Potter, J. D., McTiernan, A., & Ulrich, C. M. (2009). A yearlong exercise intervention decreases CRP among obese postmenopausal women. Medicine and Science in Sports and Exercise, 41(8), 1533–1539. http://doi.org/10.1249/MSS.0b013e31819c7feb [ Links ]
Chodzko-Zajko, W. J., Proctor, D. N., Fiatarone Singh, M. A., Minson, C. T., Nigg, C. R., Salem, G. J., & Skinner, J. S. (2009). American College of Sports Medicine position stand. Exercise and physical activity for older adults. Medicine and Science in Sports and Exercise, 41(7), 1510–1530. http://doi.org/10.1249/MSS.0b013e3181a0c95c [ Links ]
Egger, M., Davey Smith, G., Schneider, M., & Minder, C. (1997). Bias in meta-analysis detected by a simple, graphical test. BMJ (Clinical Research Ed.), 315(7109), 629–634. [ Links ]
Higgins, J. P. T., & Green, S. (Eds.). (2008). Cochrane Handbook for Systematic Reviews of Interventions. Chichester, UK: John Wiley & Sons, Inc. [ Links ]
Higgins, J. P. T., Thompson, S. G., Deeks, J. J., & Altman, D. G. (2003). Measuring inconsistency in meta-analyses. BMJ (Clinical Research Ed.), 327(7414), 557–560. http://doi.org/10.1136/bmj.327.7414.557 [ Links ]
Hong, S. (2011). Can we jog our way to a younger-looking immune system? Brain, Behavior, and Immunity, 25(8), 1519–1520. http://doi.org/10.1016/j.bbi.2011.08.002 [ Links ]
Lages, C. S., Suffia, I., Velilla, P. A., Huang, B., Warshaw, G., Hildeman, D. A., Chougnet, C. (2008). Functional regulatory T cells accumulate in aged hosts and promote chronic infectious disease reactivation. Journal of Immunology, 181(3), 1835–1848. [ Links ]
Pawelec, G. (2006). Immunity and ageing in man. Experimental Gerontology, 41(12), 1239–1242. http://doi.org/10.1016/j.exger.2006.09.005 [ Links ]
Simpson, R. J. (2011). Aging, persistent viral infections, and immunosenescence: can exercise «make space»? Exercise and Sport Sciences Reviews, 39(1), 23–33. http://doi.org/10.1097/JES.0b013e318201f39d [ Links ]
Simpson, R. J., Cosgrove, C., Chee, M. M., McFarlin, B. K., Bartlett, D. B., Spielmann, G., Shiels, P. G. (2010). Senescent phenotypes and telomere lengths of peripheral blood T-cells mobilized by acute exercise in humans. Exercise Immunology Review, 16, 40–55. [ Links ]
Simpson, R. J., Cosgrove, C., Ingram, L. A., Florida-James, G. D., Whyte, G. P., Pircher, H., & Guy, K. (2008). Senescent T-lymphocytes are mobilised into the peripheral blood compartment in young and older humans after exhaustive exercise. Brain, Behavior, and Immunity, 22(4), 544–551. http://doi.org/10.1016/j.bbi.2007.11.002 [ Links ]
Simpson, R. J., Florida-James, G. D., Cosgrove, C., Whyte, G. P., Macrae, S., Pircher, H., & Guy, K. (2007). High-intensity exercise elicits the mobilization of senescent T lymphocytes into the peripheral blood compartment in human subjects. Journal of Applied Physiology, 103(1), 396–401. http://doi.org/10.1152/japplphysiol.00007.2007 [ Links ]
Simpson, R. J., & Guy, K. (2010). Coupling aging immunity with a sedentary lifestyle: has the damage already been done?--a mini-review. Gerontology, 56(5), 449–458. http://doi.org/10.1159/000270905 [ Links ]
Simpson, R. J., Lowder, T. W., Spielmann, G., Bigley, A. B., LaVoy, E. C., & Kunz, H. (2012). Exercise and the aging immune system. Ageing Research Reviews, 11(3), 404–420. http://doi.org/10.1016/j.arr.2012.03.003 [ Links ]
Spielmann, G., McFarlin, B. K., OConnor, D. P., Smith, P. J. W., Pircher, H., & Simpson, R. J. (2011). Aerobic fitness is associated with lower proportions of senescent blood T-cells in man. Brain, Behavior, and Immunity, 25(8), 1521–1529. http://doi.org/10.1016/j.bbi.2011.07.226 [ Links ]
Verhagen, A. P., de Vet, H. C., de Bie, R. A., Kessels, A. G., Boers, M., Bouter, L. M., & Knipschild, P. G. (1998). The Delphi list: a criteria list for quality assessment of randomized clinical trials for conducting systematic reviews developed by Delphi consensus. Journal of Clinical Epidemiology, 51(12), 1235–1241. [ Links ]
Wang, J.-S., Chen, W.-L., & Weng, T.-P. (2011). Hypoxic exercise training reduces senescent T-lymphocyte subsets in blood. Brain, Behavior, and Immunity, 25(2), 270–278. http://doi.org/10.1016/j.bbi.2010.09.018 [ Links ]
Wang, J.-S., & Lin, C.-T. (2010). Systemic hypoxia promotes lymphocyte apoptosis induced by oxidative stress during moderate exercise. European Journal of Applied Physiology, 108(2), 371–382. http://doi.org/10.1007/s00421-009-1231-2 [ Links ]
Agradecimentos:
Nada a declarar
Conflito de Interesses:
Nada a declarar.
Financiamento:
Nada a declarar.
Artigo recebido a 06.01.2014; Aceite a 25.06.2014
* Autor correspondente: Faculdade de Ciências do Desporto e Educação Física - Universidade de Coimbra Pavilhão III - Santa Clara, 3040-156, Coimbra, Portugal E-mail: lucielegm@gmail.com
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}