








Serviços Personalizados
Journal
Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Ciências Agrárias
versão impressa ISSN 0871-018X
Rev. de Ciências Agrárias vol.41 no.2 Lisboa jun. 2018
https://doi.org/10.19084/RCA17136
ARTIGO
Crescimento inicial de cana-de-açúcar inoculada com fungos micorrízicos arbusculares e fósforo
Initial growth of sugarcane inoculated with arbuscular mycorrhizal fungi and phosphorus
Jolimar Antonio Schiavo*, Luciane Silveira Azevedo, Miriam Ferreira Lima, Naelmo de Souza Oliveira e Valquíria Rodrigues Lopes
Universidade Estadual de Mato Grosso do Sul, Unidade Universitária de Aquidauana, Departamento de Solos. Rodovia Aquidauana, KM 12, Zona Rural, s/no, CEP 79200-000 Aquidauana, MS, Brasil
(*E-mail: schiavo@uems.br)
RESUMO
O objetivo deste trabalho foi avaliar os efeitos da inoculação com diferentes espécies de fungos micorrízicos arbusculares (FMAs) e de doses de P (KH2PO4), sobre o crescimento e teores de N e P em plantas jovens de cana-de-açúcar. O experimento foi conduzido em estufa utilizando delineamento experimental inteiramente casualizado, em esquema fatorial 6×5+1, com 4 repetições, sendo avaliados os fatores: doses de P (0, 50, 100, 200, 400 e 800 mg kg-1), inoculação com FMAs (Claroideoglomus etunicatum, Gigaspora albida, Gigaspora margarita, Rhizophagus clarus e FMAs nativos da área de cultivo de cana-de-açúcar), e uma testemunha sem inoculação. Aos 60 e 90 dias após o plantio, na dose 50 mg kg-1 de P as plantas inoculadas com G. albida tiveram maior altura. Plantas inoculadas com G. albida, na dose 0 mg kg-1 de P, apresentaram incremento de 300 % na matéria seca da parte aérea em relação a ausência de inoculação. A produção máxima de matéria seca da parte aérea das plantas inoculadas com G. albida foi de 149 g associada a 424 mg kg-1 de P. A inoculação com G. albida favorece o crescimento inicial e conteúdo de N das plantas de cana-de-açúcar, cuja dependência e eficiência micorrízica foi de 13,6 % e 254 %, respectivamente, na dose 0 mg kg-1 de P.
Palavras-chave: micorrizas, nutrição mineral, qualidade de mudas. Saccharum officinarum L., variedade RB 535054.
ABSTRACT
The objective of this study was to evaluate the effect of inoculation with different arbuscular mycorrhizal fungi (AMF) species and P levels (KH2PO4) on growth and nitrogen (N) and P contents in plants of sugarcane. The experiment was conducted in a greenhouse using a completely randomized design in a factorial 6×5+1, with 4 replicates, the factors being evaluated: levels of P (0, 50, 100, 200, 400 and 800 mg kg-1), inoculation with AMF (Claroideoglomus etunicatum, Gigaspora albida, Gigaspora margarita, Rhizophagus clarus and indigenous AMF in the area of sugarcane cultivation) and a control without inoculation. At 60 and 90 days after planting at a level 50 mg kg-1 of P plants inoculated with G. albida had greater height. Plants inoculated with G. albida, in level 0 mg kg-1 of P, had 300 % increase in shoot dry matter compared to the absence of inoculation. The maximum production of dry matter of shoots of plants inoculated with G. albida was 149 g associated with 424 mg kg-1 of P. The inoculation with G. albida favors the initial growth and N content of the sugarcane plants, whose dependence and mycorrhizal efficiency was 13.6% and 254%, respectively, at the dose 0 mg kg-1 of P.
Keywords: mineral nutrition, mycorrhizae, Saccharum officinarum L., seedlings quality, variety RB 535054.
INTRODUÇÃO
O Brasil é o maior produtor mundial de cana-de-açúcar (Saccharum spp.), destinada principalmente à fabricação de açúcar e agroenergia, destacando-se a região Centro-Sul, a qual apresentou expansão de área cultivada de 6,5 % na safra 2014/2015 (FAOSTAT, 2014). No sistema de produção, incluindo cana planta e soca, o ciclo da cultura varia de cinco a oito anos, com a produtividade diminuindo com o passar do tempo, principalmente em função da extração dos nutrientes do solo (Datta e Kulkarni, 2012). Entre os nutrientes, o fósforo (P), pelos baixos teores presentes no solo e pela elevada adsorção dos óxidos de ferro (Fink et al., 2014), frequentes nas áreas de cultivo da cana-de-açúcar, tem sido o principal fator responsável pela diminuição da produtividade.
Em canaviais com colheita mecanizada, com manutenção da matéria orgânica e fertilidade do solo, a queda da produtividade tem sido menos acentuada, e entre outros fatores, é atribuída à grande diversidade de fungos micorrízicos arbusculares (FMAs) presentes na rizosfera da cultura (Reis et al., 1999; Ambrosano et al., 2010; Datta e Kulkarni, 2012).
Os FMAs são simbiontes biotróficos obrigatórios e estão associados com as raízes de mais de 80 % das espécies de plantas terrestres (Smith e Read, 2008). Nesta associação, o fungo depende da planta hospedeira para alimentação e reprodução e, em troca fornece fosfato e nutrientes minerais essenciais do solo para a planta hospedeira. Uma das funções dos FMAs é de transporte de P. O micélio extra-radical dos FMAs acessam o P do solo, transportando-o até as células do cortex na forma de polifosfatos, os quais após a solubilização são translocados para a planta hospedeira (Melloni et al., 2000).
Entre os reguladores da taxa de crescimento fúngico intraradicular, o P tem sido bastante estudado. De maneira geral, altas concentrações de P na planta inibem a colonização das raizes pelos FMAs, enquanto baixas concentrações favorecem a colonização intraradicular. Esse padrão é atribuído ao fato de que altas concentraçoes de P promovem menor permeabilidade da membrana e exsudação radicular, diminuindo a nutrição do FMAs; aumento da sacarose transportada via floema até as raízes inibindo o crescimento fúngico e modulação de genes específicos da simbiose através da alteração e fitohormonios nas raízes (Kiriachek et al., 2009). No entanto, este padrão de resposta tem sido contraditório, e varia de acordo com a espécie vegetal e de FMAs. Alguns trabalhos têm mostrado que elevados teores de P no solo não interferem na colonização micorrízica (Almeida, 2007; Tellechea, 2007) e nos benefícios promovidos pelos FMAs às plantas (Sawyer et al., 2003).
Na cultura da cana-de-açúcar, principalmente na fase inicial, o efeito dos FMAs é observado pelo maior crescimento das plantas. Isto é obtido, principalmente, por meio do incremento na absorção de nutrientes, em especial o P (Berbara et al., 2006). Prabudoss (2011), trabalhando com oitos isolados de FMAs, verificou que Glomus fasciculatum promoveu maior altura e diâmetro dos colmos das plantas, bem como maior colonização micorrízica e números de esporos.
No entanto, ainda não está claro se as plantas de cana-de-açúcar são capazes de selecionar FMAs eficientes quanto à estimulação do seu crescimento. Dessa forma, é importante entender como o incremento da diversidade de FMAs no solo influencia as plantas e quais combinações planta-fungo ocorrem preferencialmente e são mais eficientes. O objetivo deste trabalho foi avaliar os efeitos da inoculação com diferentes espécies de FMAs e de doses de P sobre o crescimento inicial e sobre os teores e conteúdos de nitrogênio (N) e P em plantas de cana-de-açúcar.
MATERIAL E MÉTODOS
O trabalho foi realizado em estufa da Universidade Estadual de Mato Grosso do Sul (UEMS), com delineamento experimental inteiramente casualizado em esquema fatorial 6×5+1, sendo os fatores doses de P (0, 50, 100, 200, 400 e 800 mg kg-1) inoculação com os FMAs Gigaspora margarita (Becker e Hall), Rhizophagus clarus (Nicolson e Schenck), Claroideoglomus etunicatum (Becker e Gerdemann), Gigaspora albida (Schenck e Smith) e Acaulospora scrobiculata (Gerdemann e Trappe) e uma testemunha sem inoculação, com quatro repetições.
Os FMAs G. margarita, R. clarus, C. etunicatum, G. albida são provenientes da coleção do laboratório de microbiologia da UEMS; enquanto, A. scrobiculata foi isolado da rizosfera de plantas de cana-de-açúcar em cultivos comerciais no Município de Maracaju – MS em 2014.
A multiplicação dos isolados foi realizada tendo como substrato uma mistura de solo e areia na proporção de 1:2 (v/v). O substrato foi esterilizado em autoclave por três vezes, à temperatura de 1210 C, por uma hora, e a seguir colocado em vasos com 5 dm3 de capacidade. Em cada vaso adicionou-se uma amostra de 50 g de solo contendo esporos e raízes colonizadas com os FMAs, a 3 cm da superfície. A seguir, foram semeadas sementes de Urocloa bryzantha, que tiveram a superfície esterilizada com solução a 0,5% de hipoclorito de sódio, durante 15 minutos. Após a embebição, as sementes foram lavadas com água esterilizada quatro vezes consecutivas. Os vasos foram mantidos em estufa por período de quatro a seis meses para a multiplicação dos FMAs, os quais foram utilizados como inóculo.
Para o experimento o substrato foi constituído por uma mistura 1:2 v/v de vermiculite média com o horizonte subsuperficial de um Argissolo Vermelho Distrófico (Schiavo et al., 2010), coletado no campo experimental da UEMS, unidade de Aquidauana, cuja análise química apresentou os seguintes resultados: pH (H2O) (1:2,5) =4,8; P (Mehlich-1)=3,5 mg kg-1; K=1,6 mmolcdm-3; Ca=10,0 mmolc dm-3; Mg=7,0 mmolc dm-3; Al=4,0 mmolc dm-3; H+Al=27,0 mmolc dm-3; SB=18,6 mmolc dm-3; CTC=45,6mmolc dm-3; V = 40,79 %; m= 8,7 %; M.O=13,0 g kg-1.
O substrato foi autoclavado à temperatura de 121ºC, por 1 h, e posteriormente acondicionado em vasos plásticos com capacidade de 5 L, sendo realizada a calagem para atingir o valor de saturação por bases de 60 %. Após a aplicação do calcário os vasos foram mantidos próximos à humidade da capacidade de campo por um período de 20 dias. Decorrido este período procedeu-se a aplicação das doses de P utilizando como fonte o KH2PO4. Em função das doses crescente de P, fez-se necessário equilibrar as doses de potássio, utilizando-se como fonte o KCl.
A variedade de cana-de-açúcar utilizada foi a RB 535054, coletada na Usina LDC-SEV (Louis Dreyfus Commodities - Santelisa Vale) localizada no município de Maracaju, MS. Após a coleta, os colmos foram cortados em toletes contendo um nó cada (microtoletes de 10 cm), e submetidos ao tratamento térmico em água aquecida (50 ºC durante 30 min.). Após este procedimento, em cada vaso foi plantado três microtoletes e concomitantemente colocado, 15 mL dos inoculantes de FMAs, composto pela mistura do substrato de multiplicação do FMA e as estruturas dos fungos micorrízicos (esporos, hifas e raízes com fragmentos de hifas).
Após 10 dias do plantio, com brotamento das gemas dos microtoletes, foi realizado o desbaste deixando-se duas plantas com maior vigor por vaso.
Foram realizadas medições de altura e diâmetro a altura do colo aos 30, 60 e 90 dias após o plantio (DAP) quando as plantas foram colhidas. Separou-se a parte aérea do sistema radicular e cerca de 10 gramas de raízes de cada planta foram coletados, e armazenadas em frascos de vidro e conservadas em solução aquosa de etanol 50 %. Nestas raízes, a determinação da colonização micorrízica foi realizada pelo método da interseção em placa de Petri reticulada (Giovannetti e Mosse, 1980), após a coloração das raízes com azul de metila, com adaptações ao método proposto por Grace e Stribley (1991).
A parte aérea e o restante do sistema radicular, após lavagem em água corrente, colocados em sacos de papel, foram levados em estufa de ventilação de ar forçada à 65 ºC por 72 h. Após a secagem foram determinadas a massa seca da parte aérea, raízes e estimado a massa total das plantas; bem como calculou-se segundo Plenchette et al. (1983) a dependência e eficiência micorrízica.
As duas primeiras folhas apicais de cada planta foram separadas do restante da parte aérea, moídas em moinho tipo Willey, e submetidas à digestão sulfúrica (para determinação de N) e digestão nítricoperclórica (para determinação de P); o N foi determinado pelo método de Nessler (Jackson, 1965) e o P por colorimetria, pelo método do molibdato de amónio (Malavolta et al., 1997).
Os dados obtidos foram submetidos à análise de variância, e quando significativo, as médias dos tratamentos de inoculação por fungos micorrízicos arbusculares e as doses de fósforo a comparação de médias, pelo teste de Tukey, a 5 %, e análise de regressão, respectivamente. Além de significativo para o ajuste de regressão, assumiu-se a utilização do modelo que apresentasse R2 ≥ 0,70.
RESULTADOS E DISCUSSÃO
A colonização micorrízica não apresentou ajuste de regressão em função das doses de P; e na dose 0 mg kg-1 de P, plantas inoculadas com C. etunicatum e G. albida tiveram maior colonização (82,5 %) em relação às sem inoculação (32,5 %), porém não diferiram dos demais inóculos. De forma semelhante, na dose de 50 mg kg-1 de P as plantas inoculadas com G. albida apresentaram maior colonização (100 %) em comparação às sem inoculação (60 %), mas não diferiram dos demais inóculos (Quadro 1). Nas demais doses de P, a colonização micorrízica foi elevada (70 a 100 %) e os inoculantes não diferiram em comparação às plantas sem inoculação.
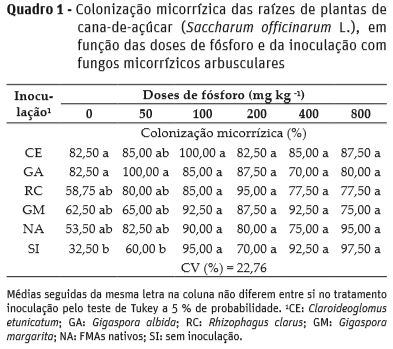
Em trabalho com seis variedades de cana-de-açúcar Jamal-Ally (2013), constatou colonização micorrízica variando de 26 a 41 %, sendo os maiores valores correlacionados à maior absorção de nutrientes. Tellechea (2007) verificou que mesmo em doses elevadas de P, plantas de cana-de-açúcar quando inoculadas com G. clarum e G. etunicatum apresentaram elevada colonização micorrízica (42 a 80 %). Segundo Almeida (2007), apenas 3 % dos genes de cana-de-açúcar manifestam sua expressão durante a micorrização, sendo a maior parte destes sem diferenciação para alto teor de P inorgânico, sugerindo que altos níveis de P podem não inibir a colonização micorrízica, corroborando com os dados deste trabalho.
No tratamento sem inoculação, constatou-se a presença de estruturas dos FMAs nas raízes (colonização acima de 32,5 %), isto pode ser devido à presença de esporos na água utilizada para irrigação das plantas, ou nos microtoletes da cana-de-açúcar; ou a propágulos de FMAs resistentes ao processo de esterilização.
A altura das plantas, em função da inoculação com os FMAs e doses de P apresentou padrão distinto nas épocas de avaliação (Figura 1). Aos 30 DAP para cada dose de P não houve diferença nas alturas das plantas entre os FMAs; e à exceção de C. etunicatum e G. albida todos os inóculos apresentaram ajuste de regressão quadrática em função das doses de P (Figura 1a).
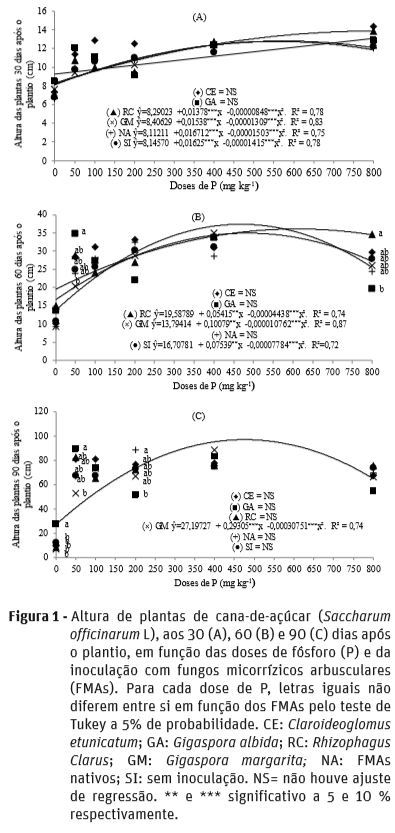
Aos 60 DAP, na dose de 50 mg kg-1 de P, plantas inoculadas com G. albida apresentaram maior altura em relação às inoculadas com G. margarita, não diferindo dos demais inóculos; enquanto na dose de 800 mg kg-1 de P, a maior altura ocorreu nas plantas inoculadas com R. clarus em relação às inoculadas com G. albida (Figura 1b). Nessa mesma época de avaliação, apenas R. clarus, G. margarita e plantas sem inoculação tiveram ajuste de regressão quadrática em função das doses de P.
Aos 90 DAP plantas inoculadas com G. albida na dose de 0 mg kg-1 de P apresentaram maior altura em relação aos demais inóculos e na dose de 50 mg kg-1 de P foram superiores em relação as inoculadas com G. margarita; enquanto na dose de 200 mg kg-1 de P a inoculação com FMAs nativos proporcionou maior altura das plantas em relação as inoculadas com G. albida. Aos 90 DAP apenas plantas inoculadas com G. margarita tiveram ajuste de regressão quadrática em função das doses de P (Figura 1c).
Quanto ao diâmetro à altura do colo, as plantas de cana-de-açúcar apresentaram pouca variação em função da inoculação com os FMAs e doses de P, com ajuste de regressão quadrática apenas para R. clarus aos 30 DAP, sendo o diâmetro máximo de 0,97 cm, associado à 576 mg kg-1 de P (Figura 2). Nesta época de avaliação, na dose de 200 mg kg-1 de P, plantas inoculadas com C. etunicatum tiveram maior diâmetro a altura do colo em comparação às inoculadas com G. albida.
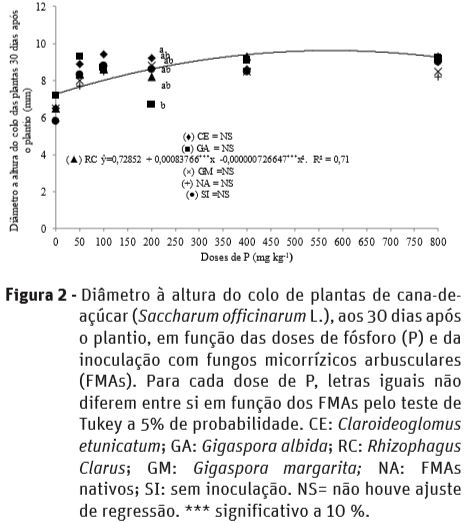
Aos 60 e 90 DAP plantas inoculadas com G. albida na dose de 200 mg kg-1 de P tiveram menor diâmetro à altura do colo em relação às sem inoculação, porém não diferiram dos demais inóculos; e na dose de 800 mg kg-1 de P, aos 60 DAP plantas inoculadas com C. etunicatum e R. clarus tiveram maior diâmetro à altura do colo em relação às inoculadas com G. albida; semelhantemente, com a aplicação de 800 mg kg-1 de P, aos 90 DAP, plantas inoculadas com G. margarita, FMAs nativos e sem inoculação, apresentaram maior diâmetro à altura do colo, em relação às inoculas com G. albida (Quadro 2).
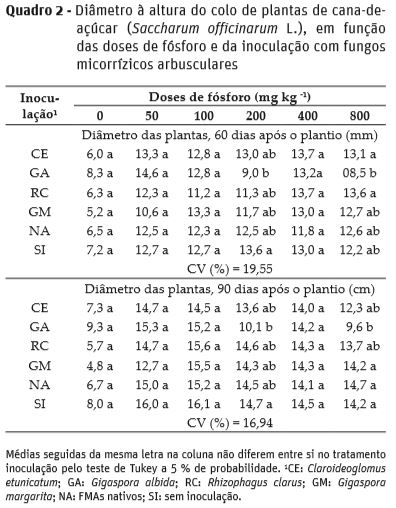
O diâmetro à altura do colo das plantas jovens e a relação altura/diâmetro à altura do colo (Campos e Uchida, 2002), são variáveis bastantes utilizadas para inferir sobre sua qualidade, apresentando boa relação com seu estabelecimento e sobrevivência a campo, às quais podem ser melhoradas com a inoculação com FMAs, fato observado neste trabalho quando procedeu-se a inoculação com G. albida, principalmente nas menores doses de P.
Plantas de cana-de-açúcar apresentaram ajuste de regressão quadrática em função das doses de P somente quando inoculadas com G. albida, com produção máxima de 149 g de massa seca da parte aérea (associada à dose de 424 mg kg-1 de P) e 175 g de massa seca total das plantas (associada à dose de 425 mg kg-1 de P), respectivamente (Figura 3a e 3b). Plantas inoculadas com G. albida na dose de 0 mg kg-1 de P tiveram maior massa seca da parte aérea em relação aos demais inóculos com incremento de 300 % em relação às sem inoculação; enquanto, na dose de 800 mg kg-1 de P as inoculadas com R. clarus é que apresentaram maior massa seca da parte aérea. De forma semelhante, a massa seca total na dose 0 mg kg-1 de P foi maior nas plantas inoculadas com G. albida em relação aos demais inóculos, com incremento de 364 % em relação às sem inoculação.
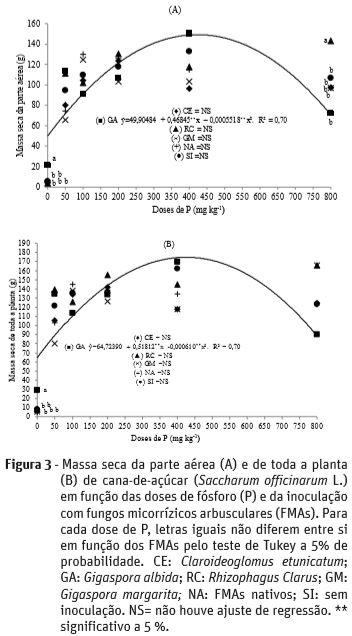
Os benefícios dos FMAs durante o ciclo vegetativo da cana-de-açúcar têm sido pouco relatados e contraditórios. Tellechea (2007), não verificou efeito dos FMAs R. clarus e G. etunicatum no acúmulo de massa seca da parte aérea e das raízes das plantas de cana-de-açúcar. No entanto, estudando o efeito de seis espécies de FMAs sobre o desenvolvimento de três variedades de cana-de-açúcar, Andreola et al. (1985), concluíram que a eficiência da inoculação pode variar de acordo com a espécie de FMAs e com a variedade da cana-de-açúcar. Adicionalmente, Reis et al. (1999) e Datta e Kulkarni (2012) observaram grande diversidade de espécies de FMAs na rizosfera de plantas de cana-de-açúcar sob diferentes condições de solo e manejo. Os fatos supracitados aliados aos resultados deste trabalho convergem para que a dependência micotrófica da cana-de-açúcar apresenta estreita relação entre as cultivares e espécies de FMAs.
Os teores de P da parte aérea não foram influenciados pela inoculação com os FMAs, e quanto às doses de P, houve ajuste de regressão linear para as plantas inoculadas com C. etunicatum, G. albida, R. clarus e sem inoculação; e quadrática para G. margarita e FMAs nativos (Figura 4a).
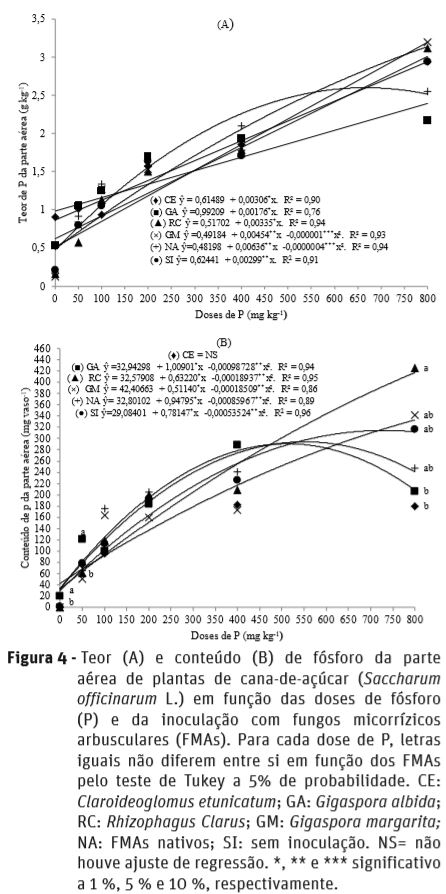
À exceção da inoculação com C. etunicatum, que não apresentou ajuste de regressão, e o tratamento sem inoculação com ajuste linear, os demais inóculos apresentaram para conteúdo de P da parte aérea, ajuste de regressão quadrática em função das doses de P (Figura 4b). Nas doses de 0 e 50 mg kg-1 de P, plantas inoculadas com G. albida tiveram maior conteúdo de P na parte aérea em relação aos demais inóculos; enquanto na dose de 800 mg kg-1 a inoculação com R. clarus proporcionou maior conteúdo de P da parte aérea em relação as inoculadas com C. etunicatum e G. albida.
Quanto ao teor e conteúdo de N na parte aérea, não houve ajuste de regressão em função das doses de P. Na dose de 800 mg kg-1 de P, plantas inoculadas com G. margarita tiveram maior teor de N na parte aérea, em relação as inoculadas com C. etunicatum, G. albida, e as sem inoculação; enquanto na dose 0 mg kg-1 de P, plantas inoculadas com G. albida apresentaram maior conteúdo de N na parte aérea em comparação aos demais inóculos (Quadro 3).
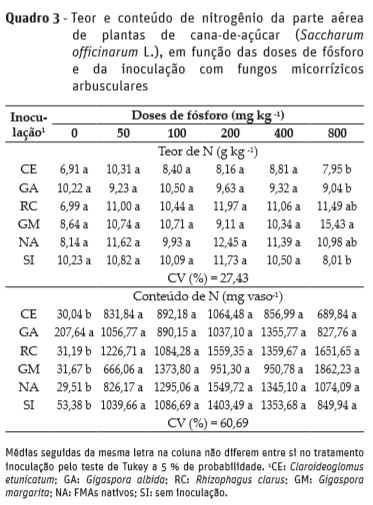
Tellechea (2007) não observou aumento no conteúdo de P da parte aérea de plantas de cana-de-açúcar inoculadas com R. clarus e G. etunicatum; porém tais conteúdos aumentaram proporcionalmente à medida que se aumentou as doses de P. Segundo este mesmo autor, as plantas também foram inoculadas com bactérias diazotróficas, promovendo alterações na anatomia do sistema radicular com aumento do número de pêlos absorventes e raízes finas, podendo ser a principal causa que minimizou os efeitos benéficos dos FMAs na absorção de P. Ainda, o mesmo autor verificou aumento nos conteúdos de N na parte aérea das plantas inoculadas apenas com G. clarum. Neste trabalho, com o incremento das doses de P no solo, observou-se que o aumento no teor de P na parte aérea foi linear, não diferindo entre os inóculos. Porém, nas doses de 0 e 50 mg kg-1 de P, plantas inoculadas com G. albida tiveram maior conteúdo de P na parte aérea. Kelly et al. (2005) verificaram que a inoculação com R. clarus incrementou o teor de P da parte aérea de plantas de cana-de-açúcar, cujo benefício para a cultura em função do FMA foi equivalente a 17 mg kg-1 de P.
Os inóculos utilizados neste experimento proporcionaram respostas distintas nas plantas de cana-de-açúcar. Destaca-se a inoculação com G. albida, que proporcionou nas menores doses de P, maior altura das plantas aos 60 e 90 DAP, maior massa seca da parte aérea e de toda a planta e maior conteúdo de P e de N da parte aérea; que, por conseguinte tiveram maior dependência e eficiência micorrízica (Quadro 4). Também, porém menos expressivo, foi verificada dependência e eficiência micorrizica em função da inoculação com R. clarus, a qual estendeu-se além das doses de 0 e 50 mg kg-1 de P (Quadro 4). Os demais inóculos propiciaram efeitos negativos de dependência e eficiência micorrizica nas plantas de cana-de-açúcar (Quadro 4), ou seja, as plantas inoculadas com FMAs apresentam menor acúmulo de massa seca, em comparação ao tratamento sem inoculação com os FMAs, mas que tiveram elevada percentagem de colonização micorrízica.
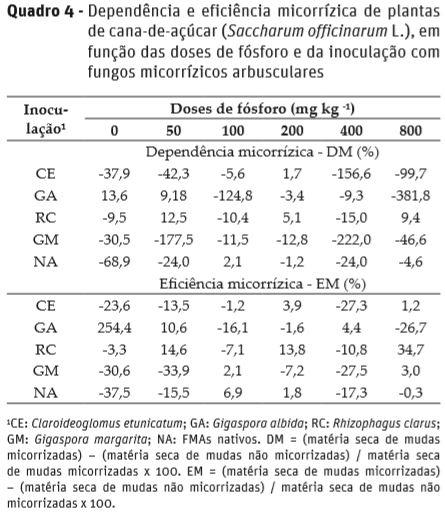
De acordo com Sawyer et al. (2003), os FMAs têm capacidade diferenciada em colonizar e promover benefícios às espécies vegetais. Ademais, o crescimento das plantas pode variar com a proporção entre micélio fúngico interno e externo. Interações negativas pode ser o resultado de um baixo crescimento externo das hifas e alta taxa de colonização radicular (Bethlenflavay et al., 1982), fato que poderia justificar neste trabalho, a baixa dependência e eficiência micorrízica dos inóculos de C. etunicatum, G. margarita e FMAs nativos. Nogueira e Cardoso (2000), avaliando produção de soja, verificaram que o FMA Gigaspora margarita proporcionou menor eficiência no estabelecimento da simbiose, fase de crescimento mais prolongada e menor produção de micélio externo ativo.
Os esporos dos FMAs são os mais importantes propágulos, cujos benefícios sobre as plantas hospedeiras dependentes da habilidade da rapidez de germinação do esporo e colonização do sistema radicular (Tommerup, 1983). Entre os vários fatores que afetam a germinação do esporo, a baixa nutrição fosfatada da planta aumenta a permeabilidade da membrana das raízes com exsudação de líquidos favorecendo a germinação com crescimento e sustentação dos FMAs durante as fases de pré e pós-infecção (Graham et al., 1981). Especificamente, espécies como a G. albida produzem múltiplos tubos germinativos, com células auxiliares, características do gênero, sendo formadas no início das ramificações hifálicas (Maia e Yano-Melo, 2001), considerado como uma estratégia de sobrevivência (Koske, 1981), podendo ser o fator responsável pela elevada colonização e aos benefícios proporcionados às plantas, conforme observado neste trabalho com este inóculo às plantas de cana-de-açúcar.
CONCLUSÕES
As espécies de fungos micorrízicos arbusculares proporcionam, na fase inicial da cana-de-açúcar, respostas diferenciadas de crescimento e acúmulo de fósforo e nitrogênio na parte aérea.
Nas doses mais baixas de fósforo (0 e 50 mg k-1 de P), as plantas de cana-de-açúcar inoculadas com Gigaspora albida tem maior altura, diâmetro à altura do colo, massa seca da parte aérea e total, teor e conteúdo de fósforo e nitrogênio da parte aérea; otimizando o uso de P do solo.
A dependência micorrízica das plantas de cana-de-açúcar apresenta estreita relação com as espécies de fungos micorrízicos arbusculares utilizados, sendo a interação Gigaspora albida versus variedade RB 535054 a mais promissora.
REFERÊNCIAS BIBLIOGRÁFICAS
Almeida, R.S. (2007) - Perfil fisiológico e da expressão de transportadores de fosfato de cana-de-açúcar (Sacharum spp.) durante a simbiose com micorrizas arbusculares. 187 p. Tese de Doutoramento em Solos e Nutrição de Plantas. Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, Brasil. [ Links ]
Ambrosano, E.J.; Azcón, R.; Cantarella, H.; Ambrosano, G.M.B.; Schammass E.A.; Muraoka, T.; Trivelin, P.C.O.; Rossi, F.; Guirado, N.; Ungaro, M.R.G. & Teramoto, J.R.S. (2010) - Crop rotation biomass and arbuscular mycorrhizal fungi effects on sugarcane yield. Scientia Agricola, vol. 67, n. 6, p. 692-701. http://dx.doi.org/10.1590/S0103-90162010000600011 [ Links ]
Andreola, F.; Cardoso, E.J.B.N. & Silveira, A.P.D. (1985) - Efeito de seis espécies de fungos micorrízicos vesiculo arbusculares sobre o desenvolvimento de três variedades de cana-de-açúcar. Tecnologia/Pesquisa STAB, vol. 4, n. 1, p. 35-37. [ Links ]
Berbara, R.L.L.; Souza, F.A. & Fonseca, H.M.A.C. (2006) - Fungos Micorrízicos Arbusculares: Muito Além da Nutrição. In: (Fernandes, M.S.; Ed.) - Nutrição Mineral de Plantas. Viçosa: Sociedade Brasileira de Ciência do Solo, vol. 1, cap. 3, pp. 53-88. [ Links ]
Bethlenflavay, G.J.; Brown, M.S. & Pacovsky, R.S. (1982) - Relationships between hosts and endophytes development in mycorrhizal soybeans. New Phytologist, vol. 90, n. 3, p. 537-543. https://doi.org/10.1111/j.1469-8137.1982.tb04486.x [ Links ]
Campos, M.A.A. & Uchida, T. (2002) - Influência do sombreamento no crescimento de mudas de três espécies amazônicas. Pesquisa Agropecuária Brasileira, vol. 37, n. 3, p. 281-288. http://dx.doi.org/10.1590/S0100-204X2002000300008 [ Links ]
Datta, P. & Kulkarni, M. (2012) - Arbuscular mycorrhizal fungal diversity in sugarcane rhizosphere in relation with soil properties. Notulae Scientia Biologicae, vol. 4, n. 1, p. 66-74. http://dx.doi.org/10.15835/nsb416567 [ Links ]
Fink, J.R.; Inda, A.V.I.; Bayer, C.; Torrent, J. & Barrón, V. (2014) - Mineralogy and phosphorus adsorption in soils of south and central-west Brazil under conventional and no-tillage systems. Acta Scientiarum Agronomy, vol. 36, n. 3, p. 379-387. http://dx.doi.org/10.4025/actasciagron.v36i3.17937 [ Links ]
FAOSTAT (2014) - Top production – World (Total). Food and Agriculture Organization of The United Nations. [cit. 2016.12.19] <http://faostat.fao.org/site/339/default.aspx> [ Links ].
Giovannetti, M. e Mosse, B. (1980) - An evaluation of techniques for measuring vesicular arbuscular mycorrizal infection in roots. New Phytologist, vol. 84, n. 3, p. 489-500. http://dx.doi.org/10.1111/j.1469-8137.1980.tb04556.x [ Links ]
Grace, C. & Stribley, D.P.A. (1991) - Safer procedure for routine staining of vesicular-arbuscular mycorrhizal fungi. Mycological Research, vol. 95, n. 10, p. 1160-1162. https://doi.org/10.1016/S0953-7562(09)80005-1 [ Links ]
Graham, J.H.; Leonard, R.T. & Menge, J.A. (1981) - Membrane-mediated decrease in root exudation responsible for phosphorous inhibition of vesicular-arbuscular mycorrhiza formation. Plant Physiology, vol. 68, n. 3, p. 548-552. https://doi.org/10.1104/pp.86.2.491 [ Links ]
Jackson, M.L. (1965) - Soil chemical analysis. New Jersey: Prentice Hall, 498p. [ Links ]
Jamal-Ally, S.F. (2013) - Arbuscular mycorrhiza and soil microbial interactions in sugarcane agriculture in Kwazulu–Natal, South Africa. 2013. 182p. Thesis - PHD - University of the Witwatersrand, Johannesburg. [ Links ]
Kelly, R.M.; Edwards, D.G.; Thompson, J.P. & Magarey, R.C. (2005) - Growth responses of sugarcane to mycorrhizal spore density and phosphorus rate. Australian Journal of Agricultural Research, vol. 56, n. 12, p. 1405-1413. https://doi.org/10.1071/AR04185 [ Links ]
Kiriachek, S.G.; Azevedo, L.C.B.; Peres, L.E.P. & Lambais, M.R. (2009) - Regulação do desenvolvimento de micorrizas arbusculares. Revista brasileira de Ciência do Solo, vol. 33, n. 1, p. 1-16. http://dx.doi.org/10.1590/S0100-06832009000100001 [ Links ]
Koske, R.E. (1981) - Gigaspora gigantea: observations on spore germination of VA-mycorrhizal fungus. Mycologia, vol. 73, n. 2, p. 73-288. [ Links ]
Maia, L.C. e Yano-Melo, A.M. (2001) - Germination and germ tube growth of the arbuscular mycorrhizal fungi Gigaspora albida indifferent substrates. Brazilian Journal of Microbiology, vol. 32, n. 4, p. 281-285. http://dx.doi.org/10.1590/S1517-83822001000400005 [ Links ]
Malavolta, E.; Vitti, G.C. & Oliveira, S.A. (1997) - Avaliação do estado nutricional das plantas: princípios e aplicações. 2. ed. Piracicaba: Potafos. Brasil.319 p. [ Links ]
Melloni, R.; Nogueira, M.A.; Freire, V.F. & Cardoso, E.J.B.N. (2000) - Fósforo adicionado e fungos micorrízicos arbusculares no crescimento e nutrição mineral de limoeiro-cravo [Citrus limonia (L.) OSBECK]. Revista brasileira de Ciência do Solo, vol. 24, n. 3, p. 767-775. http://dx.doi.org/10.1590/S0100-06832000000400009 [ Links ]
Nogueira, M.A. & Cardoso, E.J.B.N. (2000) - Produção de micélio externo por fungos micorrízicos arbusculares e crescimento da soja em função de doses de fósforo. Revista brasileira de Ciência do Solo, vol. 24, n. 2, p. 329-338. http://dx.doi.org/10.1590/S0100-06832000000200010 [ Links ]
Plenchette, C.; Fortin, J.A. & Furlan, V. (1983) - Growth responses of several plant species to mycorrhizae in a soil of moderate P-fertility. Plant and Soil, vol. 70, n. 2, p. 199-209. https://doi.org/10.1007/BF02374781 [ Links ]
Prabudoss, V. (2011) - Interaction of AM fungi and sugarcane (Saccharum officinarum L.). International Journal of Current Research, vol. 3, n. 8, p. 228-234. [ Links ]
Reis, V.M.; De Paula, M.A. & Döbereiner, J. (1999) - Ocorrência de micorrizas arbusculares e da bactéria diazotrófica Acetobacter diazotrophicus em cana-de-açúcar. Pesquisa Agropecuária Brasileira, vol.34, n. 10, p. 1933-1941. http://dx.doi.org/10.1590/S0100-204X1999001000021. [ Links ]
Sawyer, N.A.; Chambers, S.M. & Cairney, J.W.G. (2003) - Utilization of inorganic and organic phosphorus sources by isolates of Amanita muscaria and Amanita species native to temperate eastern Australia. Australian Journal of Botany, vol. 51, n. 2, p. 151-158. http://dx.doi.org/10.1071/BT02073 [ Links ]
Schiavo, J.A.; Pereira, M.G.; Miranda, L.P.M.; Dias Neto, A.H. & Fontana, A. (2010) - Caracterização e classificação de solos desenvolvidos de arenitos da formação Aquidauana-MS. Revista Brasileira de Ciência do Solo, vol. 34, n. 3, p. 881-889. http://dx.doi.org/10.1590/S0100-06832010000300029 [ Links ]
Smith, S.E. & Read, D.J. (2008) - Mycorrhizal Symbiosis. San Diego: Academic Press. 787 p. [ Links ]
Tellechea, F.R.F. (2007) - Fungos micorrízicos, bactérias diazotróficas endofíticas e fósforo no crescimento e acúmulo de nutrientes em mudas de cana-de-açúcar. 53 p. Dissertação (Mestrado em Produção Vegetal) - Universidade Estadual do Norte Fluminense Darcy Ribeiro, Campos dos Goytacazes, Brasil. [ Links ]
Tommerup, I.C. (1983) - Temperature relations of spore germination and hyphal growth of vesicular mycorrhizal fungi in soil. Transactions of the British Mycological Society, vol. 81, n. 2, p. 381-387. https://doi.org/10.1016/S0007-1536(83)80090-4 [ Links ]
AGRADECIMENTOS
À FUNDECT, chamada FUNDECT/CNPq 02/2007-PPP; ao CNPq/CAPES pelo apoio financeiro via projetos: casadinho processo 620029/20080 e casadinho/Procad processo 552377/2011-2; e a CAPES pela concessão de bolsa de estudo.
Recebido/received: 2017.06.06
Recebido em versão revista/received in revised form: 2017.10.24
Aceite/accepted: 2017.11.30